Evolution
Articles
- L'évolution du sexe
- Histoire des idées sur l'évolution:
- En attendant Darwin...
- Le débat sur l'hérédité
- Les forces évolutives
- Les Contraintes du développement

General
L'évolution du sexe...

Bien que vous ne soyez pas forcément arrivé sur cette page par le plus grand des hasards, cette dernière ne correspondra peut être pas à
vos attentes secrètes. Cependant, prenez tout de même le temps de la lire : elle peut vous
introduire au mystère le plus grand de la biologie du vingtième siècle:
Pourquoi les espèces sont elles
majoritairement sexuées?
Le rôle de cette page n'est en effet pas seulement d'augmenter le traffic sur ce site
grâce à un mot clé particulièrement prisé des internautes (mais sicela arrive, tant mieux!), mais bel et bien
de résumer un problème qui n'a pas fini de donner du fil à retordre à tout biologiste de l'évolution. En
espérant qu'elle vous éclaire sur le sujet...
Introduction : Qu'est ce que le sexe?

Les belles etamines
et le style
du Lys Martagon...
Si l'on devait s'en tenir à une définition assez large, quel est le point commun aux différentes manières dont s'exprime le sexe chez les êtres vivants? Vous avez probablement une petite idée en tête : des organes sexuels bien sûr! Fort juste, mais cela ne sera pas suffisant : non seulement leur disparité est incroyable (vous pensiez aux étamines chez les plantes ou aux cystes chez les algues je présume?), mais certains sont à peine différenciés,ou alors il faut imaginer les protiste unicellulaires, certains étant pourtant sexués, comme des organes sexuels??? Non. Insuffisant, donc.
Grain de pollen...
Si ce ne sont les organes, peut être sont-ce les gamètes me diriez vous? Oui et non, si l'on prend comme exemple la levure, cette espèce est sexuée sans pour autant produire de gamètes... Alors ce n'est toujours pas ça, mais nous nous en approchons.

Méiose male chez Aloe
Nous nous baserons donc sur cette définition relativement large : le sexe (la reproduction sexuée), c'est l'alternance d'un cycle constitué de la méiose et de la fusion de gamètes haploïdes. Il en existe d'autres, plus restreintes (probablement celle que vous aviez en tête en arrivant sur ce site) ou plus larges (définissant le sexe comme un échange d'information génétique, ce qui y inclut la recombinaison chez les bactéries), mais nous ne nous y attarderons pas trop.
<--- L'insuffisance de la réponse classique comme raison d'exister du sexe...
- 1. Le coût du sexe (coût de la méiose ou coût de la production de mâles)
- 2. Différentes hypothèses expliquant son maintien au cours de l'évolution...
- a. La fixation des mutations favorables et l'adaptation
- b. La purge des mutations délétères et le cliquet de Müller
- c. L'hypothèse de la Reine Rouge
- 3. Origine du sexe et de la méiose...
- 4. Les conséquences du sexe
- a. La naissance des genres 'femelle' et 'mâle'...
- b. La stabilité du sexe ratio 50-50
- c. La sélection sexuelle
- Conclusions
L'insuffisance de la réponse classique comme raison d'exister du sexe...

Nymphéa...
La vision classique de la raison d'être du sexe est la suivante : le sexe, c'est bien, cela permet de brasser et de recombiner l'information génétique et cela permet aux descendants de s'adapter. Le rôle du sexe dans l'apparition de nouvelles combinaisons génétiques est effectivement important, et l'idée qu'il s'agit à la fois de son rôle originel et de la raison de son maintien est ancré dans les esprits. Si l'on devait retracer l'histoire de cette proposition, la piste nous mène à un grand biologiste de la fin du 19ème, Auguste Weisman (connu également pour avoir distingué la lignée des cellules sexuelles, le germen, de la lignée somatique), pour qui le sexe était même plus que ça : il le considérait directement comme créateur de la variation dans la nature. Ce rôle sera finalement délégué à la mutation, mais l'importance indéniable du sexe dans la recombinaison est probablement à l'origine de cette conception classique du sexe. Cependant, cet avantage du sexe lié à la recombinaison n'est qu'un avantage sur le long terme. A court terme, la recombinaison est assez peu efficace, et nécessite un effectif important. Or comme nous allons le voir, à cette échelle, d'autres stratégies (asexuées) sont plus efficaces adaptativement et peuvent déstabiliser la reproduction sexuée et la remplacer.. Les deux questions fondamentales du sexe demeurent : pourquoi est-il apparu, et pourquoi est-il toujours là?
La définition classique, et encore assez répandue, selon laquelle le sexe existe parce qu'il créée de la diversité n'explique ni pourquoi le sexe est apparu au cours de l'évolution, ni comment cet avantage de diversité ne permet vraiment son maintien face a des stratégies asexuées, comme nous allons le voir...
<--1. Le coût du sexe (coût de la méiose ou coût de la production de mâles)
Il est difficile de comprendre pourquoi la réponse classique de la raison d'exister du sexe est très insuffisante pour expliquer l'abondance de la sexualité comme mode de reproduction chez les espèces vivantes, avant d'avoir fait le petit exercice suivant :
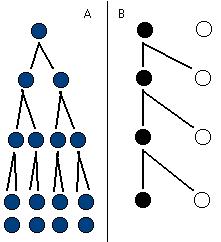
(A) Parthénogenèse > sexualité (B)
Prenons, chez une espèce hypothétique, deux femelles. L'une d'elle se reproduit sexuellement (B), l'autre se reproduit par parthénogénèse (A) (elle se reproduit sans sexualité et produit des enfants qui sont des répliques génétiques d'elle même --des clones). Chacune de ses femelles produit en moyenne deux descendants. La femelle sexuée se reproduit sexuellement, elle a donc besoin d'un mâle, et parmi ses descendants, la moitié seront du sexe mâle. La femelle parthénogénétique, elle, produit deux descendants, forcément femelles et, comme elle, parthénogénétiques. Jusque là, pas de problème. Mais à la génération suivante, seule la femelle peut produire deux descendants chez les individus sexués, tandis que nos deux femelles parthénogénétiques peuvent chacune avoir deux descendants, ce qui fait quatre femelles parthénogénétiques comme petits-enfants pour notre première femelle parthénogénétique, tandis que notre première femelle sexuée n'a que deux femelles chez ses petits enfants (le mâle se reproduit aussi). Au bout de n générations, une femelle parthénogénétique aura une descendance 2 exposant n fois supérieure à celle d'une femelle sexuée: c'est à dire 1024 fois plus en l'espace de dix générations! Il peut donc suffire d'une seule femelle parthénogénétique pour surpasser une population entièrement sexuée en quelques générations... C'est ce que l'on appelle le coût de la production de mâles, qui est une perte assez sévère pour une population sexuée : elle se reproduit en moyenne deux fois moins bien qu'une population asexuée.
Ce coût du sexe est également parfois appelé coût de la méiose. On peut résumer cette idée de la manière suivante : un gène présent chez une femelle sexuée a seulement une chance sur deux d'être transmis à sa descendance (il doit laisser sa place à un gène issu d'un individu mâle). Pour que le sexe se maintienne face à l'invasion d'une stratégie asexuée, il faut que l'avantage de la sexualité soit au moins deux fois plus grand.
Or un avantage du simple au double est quelquechose d'énorme, en biologie. Si la raison d'être du sexe est de générer de la diversité, quel est l'avantage immédiat d'un individu de mélanger la moitié de ses gènes avec n'importe quel autre individu pour produire une diversité dont on ne sait même pas si elle sera avantageuse? L'autre moitié peut très bien comporter des mutations délétères, et ça, ça peut difficilement faire mieux...

Couple d'Aretaon asperrimus
La vision classique insiste donc surtout sur un avantage populationnel de la reproduction sexuée, mais un avantage hasardeux, et très lent à produire des combinaisons génétiques avantageuses. Bref, un avantage, oui, mais à long terme, et qui n'explique donc pas du tout le sexe est apparu et s'est maintenu à son origine.
En guise d'illustration ci à droite, un couple du phasme Aretaon asperrimus. Les phasmes ont ceci d'étonnant que les femelles sont dites parthénogénétiques facultatives : si elles trouvent un mâle, elles se reproduisent de façon sexuée. Si elles n'en trouvent pas, elles se reproduisent par parthénogénèse. Peut être une solution pas si bête! ;-)
La stratégie de reproduction sexuée, pour pouvoir se maintenir face à des stratégies de reproduction asexuée, doit donc surmonter un désavantage du simple au double, lié au coût de la production de mâles. Ce désavantage est énorme pour un processus biologique...
<--2. Différentes hypothèses expliquant son maintien au cours de l'évolution...
De nombreuses hypothèses ont été émises pour rendre compte du maintien du sexe dans ce contexte. Un avantage doit certainement exister, étant donné la représentation si forte de la sexualité dans le monde vivant. Chacune d'elle reflète une possibilité parmi les autres, et à l'heure actuelle aucune ne semble vraiment privilégiée par rapport aux autres. Il est également possible que la raison ultime de la prépondérance du sexe soit une mosaïque de ces nombreuse hypothèses.
a. La fixation des mutations favorables et l'adaptation
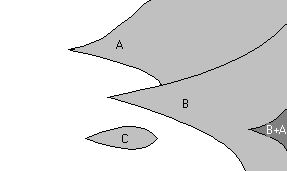
Dynamique des mutations bénéfiques
dans une population asexuée.
Un des avantages du sexe réside dans le fait que dans les population asexuées, l'adaptation dépends du taux de mutations favorables, et qu'il est très improbable que ces mutations apparaissent simultanément dans une lignée. Si de telles mutations apparaissent dans des lignées indépendantes, elles sont condamnées à se juxtaposer dans la population. Le temps nécessaire à l'apparition de ces mutations au sein de la même lignée est donc très long...
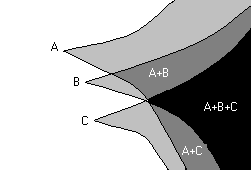
Dynamique des mutations bénéfiques
dans une population sexuée.
En revanche, si de telles mutations apparaissent dans une population se reproduisant de manière sexuée, elles pourront rapidement être cumulées au sein d'individus porteurs par le fait de la redistribution des chromosomes (ségrégation) voire entre chromosomes (recombinaison) liées à la méiose. De fait, les combinaisons avantageuses ne sont plus seulement dépendantes du taux de mutation (via la lente apparition cumulée au cours du temps), mais également de la redistribution et des nouvelles combinaisons générées grâce à la reproduction sexuée. Mais ce scénario n'est pas uniquement une version de l'explication classique, car elle va plus loin : du fait de la rapidité accrue de l'assortiment des mutations favorables, la reproduction sexuée peut être avantagée parce qu'elle accélère l'adaptation par rapport à la reproduction asexuée.
Il est parfaitement possible que, considérant l'ensemble des mutations favorables potentielles, le gain de la sexualité soit supérieur à son coût (paragraphe précédent). Cependant, cet argument dépends du taux de mutations favorables, qui est en général considéré comme plutôt faible (mais non nul!). Cela reste donc un avantage dont le bénéfice apparaît plutôt sur le long terme, l'adaptation étant un phénomène progressif. De plus, si l'argument paraît expliquer correctement le maintien du sexe, il ne résoud pas la question de son origine...
<--b. La purge des mutations délétères et le cliquet de Müller
L'hypothèse précédente dépendait de l'apparition de mutations favorables, qui sont moins fréquentes que les mutations défavorables (délétères). Voici donc une seconde hypothèse basée sur les conséquences génétiques des mutations, et qui est prend le contrepied de la précédente. Au cours du temps, des mutations délétères apparaissent inexorablement. Un organisme se reproduisant de façon asexuée ne peut donc que cumuler ces mutations : les descendants ont au mieux au moins autant de mutations que leurs parents. Cette situation rend la reproduction asexuée désavantageuse sur le long terme. Elle porte le nom de cliquet de Müller : la composition du génome qui se reproduit par voie asexuée accumule inexorablement des mutations de façon irréversible. Les organismes à reproduction sexuée, au contraire, peuvent échapper à cette acccumulation de mutations délétères : ségégration des chromosomes et recombinaison peuvent réintroduire des descendants qui possèdent moins de mutations à effets délétères que leurs parents.
c. L'hypothèse de la Reine Rouge

Alice et la reine rouge.
L'hypothèse de la Reine Rouge, pour expliquer le maintien du sexe, pourrait s'apparenter à la première hypothèse (le sexe permet aux organismes de s'adapter mieux), mais elle en diffère par son aspect dynamique, et l'adaptation qui la concerne est pour ainsi dire facultative (et s'établie sur un coût qui aurait pu être évité autrement). Venons en aux faits : l'hypothèse de la Reine Rouge fait référence au roman de Lewis Caroll ("A travers le miroir"), lorsque Alice passée au pays des merveilles, doit suivre ce conseil de la Reine et courir sans s'arrêter pour pouvoir rester à la même place. En terme biologiques, l'analogie est en effet frappante : même si l'environnement est en apparence constant, les espèces doivent sans cesse s'adapter à leur environnement biotique (parasites, maladies). En effet, les parasites s'adaptent aux défenses de leurs hôtes, qui doivent en retour trouver de nouvelles solutions pour contourner les attaques de ces parasites et ainsi de suite. Dans cette course aux armements, sans cesse renouvelée, le sexe apparaît comme une stratégie qui permet une réponse rapide aux fluctuations et à la dynamique des populations de parasites.
Des hypothèses purement génétiques basées sur la combinaison des mutations favorables, dont peuvent bénéficier les organismes à reproduction sexuée, ainsi que sur l'accumulation des mutations délétères, auxquelles les descendants issus de reproduction sexuée peuvent également échapper, pourraient expliquer le maintien du sexe en dépit d'un désavantage à court terme. Ces hypothèses peuvent prendre plus de force en les incluant dans un cadre écologique, celui de l'adaptation à l'environnement, et notamment avec l'hypothèse de la Reine Rouge qui concerne plus spécifiquement l'environnement biologique et les interactions entre espèces et notamment les interactions hôtes-parasites. Les exemples docmentés de façon empiriques sont ici trop nombreux pour être abordés en détail... Cependant, s'ils apportent tous des précisions sur le maintien du sexe, ils ne donnent aucune idée de pourquoi le sexe peut bien être apparu...
<--3. Origine du sexe et de la méiose...
L'origine du sexe peut nous donner des indications précieuses sur les raisons de son apparition, et donc de son avantage à cours terme dans cette forme originelle. Une hypothèse nouvelle sur l'utilité de la méiose, et donc de l'existence du sexe a été formulée lorsqu'il a été découvert qu'au cours de ce processus, d'importants mécanismes de correction et de réparation de l'ADN étaient activé, dont certains sont partagés avec des organismes asexués (selon la définition que nous avons adopté en début de page, à savoir caractérisés par l'existence d'une méiose au cours de leur cycle de vie) : les bactéries. Il apparaît donc qu'une des hypothèse possible de l'apparition de la méiose soit liée à la réparation de l'ADN, ce qui a pour conséquence une diminution de la variabilité. Cette hypothèse va donc dans le sens contraire de l'explication classique de l'existence du sexe. Cependant, ces mécanismes peuvent avoir évolué pour s'exprimer à ce moment particulier du cycle de vie d'une grande partie des organismes, simplement parce que l'occasion est idéale pour le faire à ce moment là : l'ADN y est moins "actif" (l'expression des gènes y est en majorité réprimée et l'ADN est condensé et sous forme de chromosomes). Cependant, la focalisation des mécanismes de réparation à ce stade de la vie des organismes a dû être mobilisée relativement tôt après l'évolution de la méiose et ce point ne peut être négligé...
<--4. Les conséquences du sexe
a. La naissance des genres 'femelle' et 'mâle'...

Dianthus sylvestris,
fleur hermaphrodite
et fleur femelle.
Quoiqu'il en soit, notre définition du sexe est très large. Certains organismes ont au cours de leur cycle de vie un passage par la méiose, puis produisent des gamètes qui fusionneront (ils sont parfois produits d'ailleurs un certain temps après la méiose elle-même, par exemple chez les plantes, sans pour autant que cette sexualité soit celle qu'on imagine. En particulier, le sexe peut exister sans qu'un genre sexuel (mâle ou femelle) n'existe. Le plus surprenant cependant, c'est que la majorité des êtres vivant sexués ont cette spécialisation en genres différents. Cette distinction repose par construction sur le type de gamètes produits : le genre mâle est défini par la production de gamètes nombreux, petits, et en général mobiles (ce critère étant le moins fort), tandis que le genre femelle est définit par des gamètes peu ou moins nombreux, volumineux et riches en réserves, et en général immobiles. Cette distinction caractérise les organes qui produisent les gamètes, car de nombreux organismes sont hermaphrodites (plantes à fleur par exemple). L'oeillet illustré ci contre par exemple, Dianthus sylvestris, montre deux types de fleurs différentes : la grande est une fleur hermaphrodite en phase mâle, tandis que la petite est une fleur femelle. Cette espèce est en réalité dite gynodioïque, c'est à dire qu'elle est composée d'individus ne présentant soit que des fleurs femelles, soit que des fleurs hermaphrodites (d'abord mâles puis femelles), ce qui est en général considéré comme une étape transitoire vers la production de sexes mâles et femelles sur des individus distincts.
Cette généralisation des genres sexuels aux espèces qui pratiquent le sexe est assez extraordinaire. Deux phénomènes sont effectivement très surprenants : d'une part, que lorsque le sexe existe, il évolue vers une différenciation en deux genres sexuels (en fait, comme nous allons le voir, deux stratégies reproductives différentes, qui se retrouvent mêmes à des niveaux divers) ; et d'autre part que ces deux options soient équilibrées : en général, il y a autant de mâles que de femelles. Ces généralités nous sont d'ailleurs si naturelles qu'elles nous semblent triviales, à tel point que s'interroger sur leur raison d'être peut même paraître étrange : pourquoi se demander pourquoi il n'y a que deux sexes, et pourquoi sont ils répartis en proportions égales chez les espèces sexuées?

Investissement de
la fonction maternelle
dans les descendants.

La naissance des genres, c'est à dire la production de gamètes différenciés en gamètes mâles ou femelles caractérisés par leurs différences en taille et en nombre produits, se comprend
mieux si l'on imagine son évolution à partir d'un état ancestral pour lequel les gamètes sont identiques (et que l'on appelle isogamie... cet état est encore rencontré dans certains organismes ). On imagine bien que fournir
un peu plus de nutriments et de réserves dans ses gamètes est avantageux et contribue à augmenter les chances de survivre à sa descendance ainsi mieux pourvue. Cette stratégie (qui débouchera par la suite sur la production de gamètes
femelles, plus volumineux), trouve une explication simple et intuitive dans le cadre du paradigme darwinien par une augmentation progressive des réserves allouées aux gamètes : les gamètes les mieux pourvus en réserves nutritives
ont autant de chance en plus de survivre.
Maintenant, comment expliquer l'évolution vers une stratégie "mâle", c'est à dire une production de gamètes avec peu de réserves? Si la réponse est maintenant un peu plus subtile, elle n'en demeure
pas moins assez simple : s'il existe dans la population des gamètes un peu plus gros que les autres, un gamète ayant autant moins de réserve a autant de chance de survivre s'il féconde ce type de gamète que si les deux gamètes
apportaient chacun autant de réserves. Il est vrai que cet absence de désavantage ne fonctionne que si cette situation se produit, et que le zygote résultant de la fécondation aura moins de chance de survivre si le gamète 'mâle' féconde
un autre gamète normalement constitué en réserves. Mais regardons un peu ce qui se passe pour l'individu qui produit des gamètes avec moins de réserves (stratégie mâle) : puisque ses gamètes sont moins coûteux en énergie, il peut en
produire plus. S'il produit plus de gamètes, il a plus de chance que les autres d'assurer une descendance. Il y a donc là bel et bien une compensation par rapport au risque de fournir moins de réserves au futur descendant...
Tout tient
dans cet étroit équilibre à trouver entre produire plus de gamètes, ou leur fournir plus de réserves pour le futur descendant... La stratégie mâle correspond à une stratégie 'tricheuse', dans la mesure où elle est avantageuse d'abord
à la reproduction de l'individu mâle plutôt qu'à la survie du descendant, et celà au détriment du partenaire associé -la femelle, qui investit plus dans le gamète. On peut également remarquer que l'investissement dans le descendant lié à la fonction
femelle va bien au delà de l'investissement dans les gamètes: les plantes l'illustrent bien, puisque les structures femelles continuent à apporter des réserves aux graines, ainsi qu'à des structures aussi coûteuses que les fruits afin d'assurer leur dispersion...
Le moins qu'on puisse dire, c'est que cette évolution vers la dissociation
de ces deux stratégies de reproduction s'est produite plusieurs fois de façon indépendante, et on peut encore observer des groupes chez lesquels cette dissociation ne s'est pas produite, notamment chez certaines algues...
L'apparition de la reproduction sexuée n'a donc pas que des conséquences purement génétiques de réarrangement de la variabilité génétique. L'une de ses conséquences les plus importantes est ausi la naissance de la spécialisation des stratégies de reproduction en deux options fondamentalement opposées et qui caractérisent les genres sexuels à différents niveaux: d'une part une stratégie "femelle", basée sur l'investissement reproducteur dans les descendants qui leur assure un avantage pour la survie ; et d'autre part une stratégie "mâle", qui profite de l'investissement lié à la fonction femelle et 'préfère' faire le pari d'investir dans la quantité (plus de gamètes produits mais avec moins de réserves), donc une stratégie du risque plutôt que de l'assurance reproductive. Ces deux stratégies sont effectivement deux voies différentes de l'optimisation de la reproduction individuelle: donner plus de chances à ces descendants, ou tenter d'avoir plus de descendants... L'existence et l'apparition récurrente de ces stratégies leur donnent un généralisme assez stupéfiant du point de vue scientifique.
<--b. La stabilité du sexe ratio 50-50
Un deuxième aspect assez étonnant de l'évolution du sexe est que non seulement il donne presqu'invariablement naissance à des genres (au moins du point de vue de la production des gamètes, c'est à dire des organes sexuels puisque de nombreuses espèces, notamment chez les plantes sont hermaphrodites), mais encore la répartition de ces genres s'équilibre de façon équiprobable: la quasi majorité des espèces sexuées ont autant de femelles que de mâles. C'est d'ailleurs assez étonnant, puisque nous avons vu que le désavantage de la reproduction sexuée est liée au coût de la production des mâles.
En fait, le mécanisme qui intervient ici est assez simple: il s'agit de l'avantage du rare. On peut le résumer rapidement de la façon suivante: si un des genres est plus rare que l'autre, le 'sexe' majoritaire est en compétition pour l'accès à la reproduction. Ainsi, les individus du sexe le plus rare tendent à se reproduire plus (relativement à ceux de l'autre sexe), et les gènes qui orientent vers la production du genre rare seront plus représentés à la génération suivante, rétablissant ainsi l'équilibre jusqu'à obtenir une répartition de moitié-moitié.
En fait, l'explication réelle est un peu plus subtile. En effet, elle n'explique qu'imparfaitement la raison de cet équilibre: les individus de sexe mâle produisent plus de gamètes qu'il n'en est nécessaire, et si leur nombre diminue, on ne fait que diminuer leur degré de compétition pour la reproduction. Les espèces qui présenteraient moins de mâles que la moitié de leur effectif ne souffriraient probablement pas d'une diminution de leur pouvoir reproducteur... Ce premier argument n'explique donc efficacement le rééquilibrage du ratio 50-50 que lorsque les individus femelles deviennent minoritaires.
Mais en reconsidérant la question du point de vue des grand-parents, le succès reproducteur optimal reste celui de produire des enfants en sexe ratio équilibré, car si les mâles sont plus rares, alors leur participation à la reproduction est plus importante: en résumé, s'il y a moins de mâles, tout gène diminuant leur rareté (en augmentant la proportion de mâles dans la descendance) permet d'avoir plus de descendants si l'on considère le nombre de petits enfants. L'avantage reproductif individuel est donc bien de tendre à produire un sexe ratio de 50-50...
Une deuxième conséquence non négligeable du sexe est que lorsque les genres sont différenciés, ils tendent à être présent en proportion moitié-moitié, et cela est la simple conséquence de l'optimisation du succès reproducteur individuel... Autrement dit, dés qu'il y a du sexe, le "coût de la production des males" est inévitable et fait peser un désavantage du simple au double à la stratégie de reproduction sexuée par rapport à celle de la reproduction asexuée. De fait, la reproduction sexuée est une stratégie instable. C'est pourquoi le fait que la grande majorité des espèces vivantes soient sexuées reste un grand mystère biologique...
<--c. La sélection sexuelle

Cardinal.
Il serait injuste de s'étendre sur l'évolution du sexe sans prendre en compte également une autre de ses conséquences majeures: la possibilité de sélection sexuelle. En effet, certaines caractéristiques des organismes, en particulier celles qui s'expriment dans un sexe particulier, peuvent difficilement s'expliquer par un avantage lié à la survie des individus qui en sont porteurs. Les meilleurs exemples de cela sont les couleurs vives où les parures extravagantes que l'on rencontre chez de nombreux mâles chez les oiseaux (comme le mâle du Cardinal illustré ci à gauche, paré d'un plumage rouge vif qui ne le prédispose pas à gagner les parties de cache-cache, ou encore comme la robe également bien coloré du paon mâle ci dessous à droite).
En fait, cela s'explique par la sélection sexuelle, qui est donc une force évolutive suffisamment forte pour contrecarrer les effets de la sélection naturelle et prodiguer certaines d'espèces d'apparats plutôt désavantageux. On imagine effectivement mal comment avoir des couleurs vives peut protéger les oiseaux de leurs prédateurs: ils n'en sont que plus visibles.

Paon mâle.

Paon femelle.
Il s'agit d'une caractéristique de la sélection sexuelle: les femelles ont des préférences pour choisir leur partenaire, et ceux qui se rapprochent le plus de ces préférences ont plus de chance de se reproduire, propageant ainsi des gènes résultant en des caractéristiques saugrenues (que l'on pense aux plumes de la queue des paons!). En fait, cela permet aussi aux femelles, si les mâles en questions sont toujours vivant au moment de la reproduction, de s'assurer de la qualité de leur partenaire, et donc des chances de survie de leurs descendants... Chez les merles, c'est la couleur du bec qui est un indicateur de résistance aux parasites, qui est favorablement séectionné. Chez les hirondelles, il s'agit de la longueur et de la symmétrie de la queue qui est l'élément du choix... Et ainsi de suite.
Le sexe, puisqu'il existe, a enfin une troisième conséquence également très importante: il permet l'émergence d'une nouvelle force évolutive, en plus du trio sélection naturelle/mutation/dérive génétique: celle de la sélection sexuelle. Charles Darwin l'avait bien perçu, mieux que son contemporain et codécouvreur de la sélection naturelle Russel Wallace, point sur lequel les deux hommes étaient en désaccord. Mais il s'agit déjà d'un autre sujet...
<--Conclusions

Syrphe butinant.
La méiose semble donc bien être apparue tout d'abord afin de réparer l'ADN. Puis le cycle a évolué d'une endoréplication vers la fusion de deux cellules haploïdes, donnant naissance au sexe tel que nous le concevons aujourd'hui. A partir de là, nous sommes devant un paradoxe: la reproduction sexuée est désavantagée par rapport aux stratégies de reproduction asexuée à court terme. A long terme, la reproduction sexuée reprend ses avantages, car elle permet une accélération de l'innovation en réassociant des mutations bénéfiques, tandis qu'elle purge également rapidement les mutations délétères. Mais cet avantage à long terme ne peut prévenir l'invasion par des stratégies asexuées. Pourtant la reproduction sexuée reste une des options les plus abondantes chez les espèces modernes.
On observe également que la plupart des espèces qui se reproduisent de façon asexuée, à certaines exceptions près comme celle des Rotifères bdelloïdes dont certaines sont asexuées depuis plusieurs millions d'années, sont des espèces très récentes. Il semble donc bien qu'il y ait un inconvénient à la reproduction asexuée qui se manifeste par une faible probabilité pour une espèce asexuée de survivre à longue échéance.

Longicornes.
La vision consensuelle et actuelle de la question du sexe est donc que les espèces qui n'ont pas su s'en passer, pour une raison ou pour une autre (les stratégies asexuées n'ont pas pu apparaître ou n'ont pas pu remplacer la stratégie sexuée pour une raison spécifique) sont finalement celles qui ont le plus de chance de survivre bien plus tard, du fait de l'avantage à long terme que finit par procurer le sexe. Les raisons qui font qu'une stratégie asexuée n'arrive pas à envahir une espèce peuvent être nombreuses, et n'ont souvent rien à voir cependant avec l'avantage à long terme du sexe. Chez certains pucerons, cette raison est qu'il est impératif de pouvoir hiverner sous forme d'oeufs, et que seuls les pucerons sexués pondent des oeufs (les femelles parthénogénétiques donnent directement naissance à des petits pucerons, et bien qu'elles finissent par être plus fréquentes que les formes sexuées à la belle saison, ces dernières reprennent le dessus en hivers). A chaque espèce sexuée correspondra donc une raison particulière expliquant pourquoi les stratégies asexuées ne l'ont pas envahi... L'évolution du sexe, c'est donc bel et bien le conte des milles et une nuits!
<--